Eloy Rodríguez Freire. Curs 2009-2010

El paper
El paper i els productes relacionats amb ell s’elaboren a partir de fibres de cel·lulosa presents a les plantes. Aquestes fibres poden provenir de diferents vegetals: cotó, fusta, palla de cereals, etc. Encara que la majoria de la producció de paper prové de la fusta (95%).
Existeixen diferents tipus de paper al mercat segons el tipus de fusta, acabament, ús i gramatge.
Fabricació de paper
El fonament bàsic per la fabricació de paper es trossejar el material vegetal amb aigua per formar una suspensió de fibres i després fer làmines de fibres superposades sobre una base porosa per poder filtrar l’aigua. Els papers especials com satinats i estucats es sotmeten a tractaments addicionals.
A la suspensió de fibres li diem pasta de paper o polpa que serà la matèria prima per fer els diferents tipus de paper. Existeixen dos mètodes químics d’obtenir aquesta pasta durant l’etapa de cocció: mètode alcalí o Kraft i mètode amb sulfits. Els dos treballen amb compostos del sofre però donen una pasta de paper amb propietats més adients segons el paper final que volem.
El mètode Kraft dóna una pasta marró més consistent per fer papers d’embalatge mentre que el mètode de sulfits fa una pasta de paper més dèbil i clar. Els dos processos permeten reciclatge dels productes químics però emeten gasos tòxics.
Passos en la fabricació de paper:


VÍDEOS dels pasos en la producció de paper:
La fabricación del papel
Cargado por mactiste. - Descubre más videos de ciencia y hi tech.
 Enzims implicats en la indústria del paper
Enzims implicats en la indústria del paper
Es necessita una gran quantitat de processos químics per convertir els arbres en paper blanc. La indústria paperera utilitza clors per blanquejar la pasta de paper i com a resultat s’obtenen compostos orgànics clorats i subproductes amb problemes de toxicitat per l’ecosistema.
Durant el procés de cocció de la pasta de paper també s’empren sulfats que són tòxics.
Els enzims poden ajudar als fabricants de paper a reduir l’ús de productes químics perjudicials com clors i sulfats. D’aquesta manera els costos en substàncies químiques i el seu posterior tractament es redueixen de manera considerable així com l’impacte mediambiental.
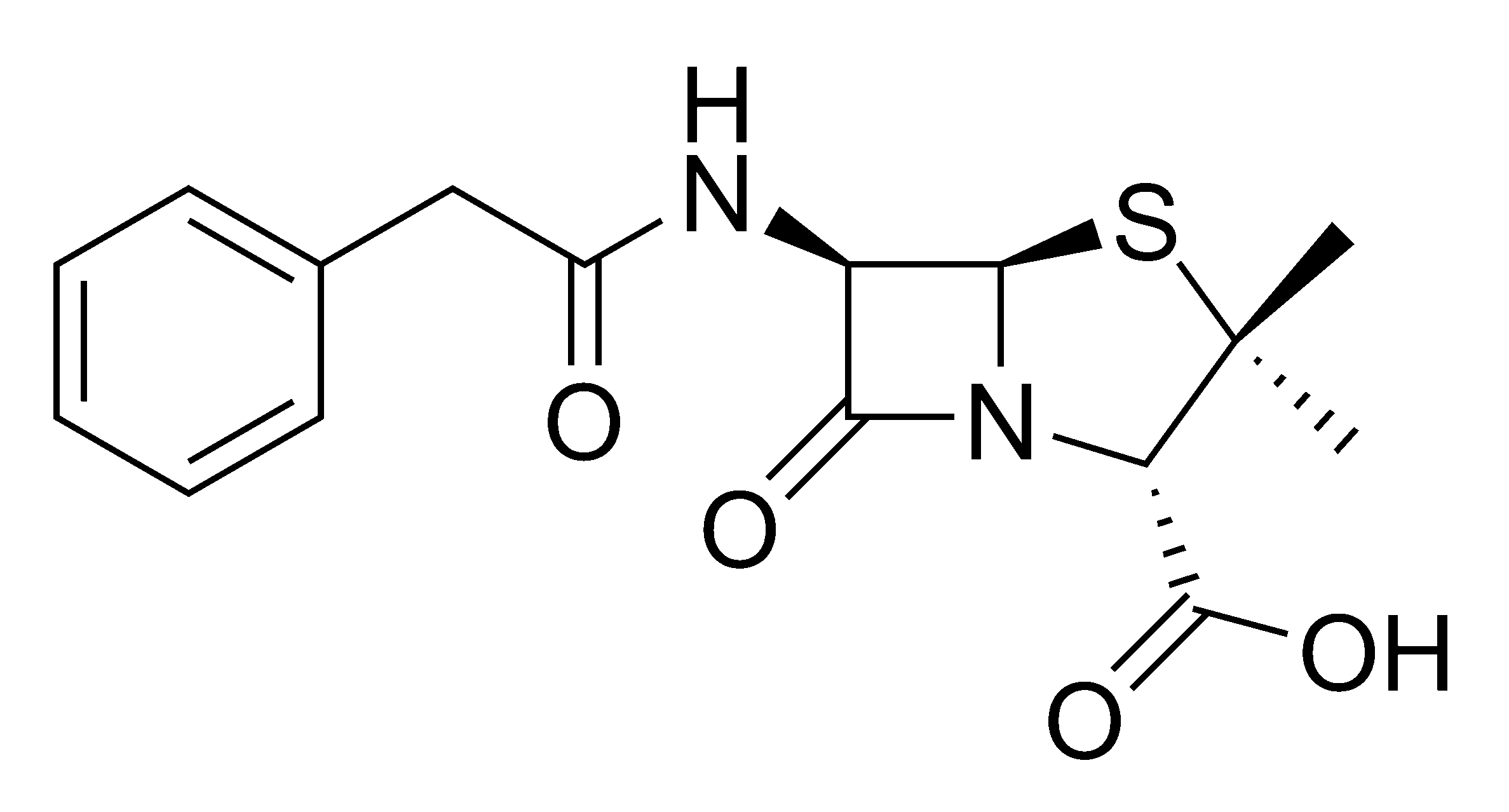
Enzims com les hemicel·luloses i la xilanasa poden millorar la decoloració reduint dràsticament l’ús de clor. El tractament enzimàtic sobre la matriu de la pasta de paper permet una millor penetració dels productes químics per blanquejar i una millor extracció al rentar la lignina i compostos associats de color marró.
Composició de la fusta per fabricar paper


Millores amb enzims a la indústria paperera en cada etapa:
Blanqueig o Bleaching
L’eliminació de la lignina de la pasta química es l’etapa de “bleaching”, es necessària per raons estètiques i per la millora de les propietats del paper. Actualment per blanquejar el paper de Kraft s’utilitzen grans quantitats de clor o derivats orgànics del clor, substàncies tòxiques, mutagèniques, persistents i bio acumulatives que poden causar nombroses alteracions perjudicials als ecosistemes biològics.
Les diferents opcions de decoloració en aquest procés son: ampliar la cocció, des-lignificació per oxigen, per diòxid de clor, la utilització de peròxid de clor, clor o ozó. La majoria d’aquestes opcions, però impliquen inversions de capital i subproductes nocius. Els enzims ofereixen una reducció de l’ús del clor i altres compostos, fan que el procés sigui més senzill i s’obté un blanqueig superior en comparació amb els mètodes actuals. A més son una opció més econòmica i ecològica per la indústria del paper.

Pel bleaching enzimàtic s’han estudiat dues estratègies possibles: hemicel·luloses i enzims lignolítics.

Thapar Corporate Research & Development Centre, India
Modificació de fibres o fibrilació:
Modificant fibres s’aconsegueix disminuir el consum d’energia a la fase tèrmica i mecànica de la producció de polpa. El tractament enzimàtic es produeix a la superfície externa de la fibra, dissolent una petita quantitat de fibra i augmentant la hidratació.
Si es combina amb un pretractament amb agents alcalins, l’acció enzimàtica redueix de manera important l’energia al procés mecànic per refinar la polpa de paper.
Els microorganismes per obtenir els enzims necessaris son Bacillus i algunes espècies d’Aspergillus.
Solubilització de la pasta:
Pastes que es dissolen per produir materials cel·lulòsics com acetats, cel·lofanes i raions. Es fa derivatitzant i solubilitzant amb alta puresa, encara no hi ha enzims específics.
Destintació o “Deinking”:
La destintació enzimàtica converteix les fibres secundàries (fibres disgregades de la pasta de paper) en productes de qualitat. Resulta un mètode més econòmic i eficaç per tractar residus de paper de reciclatge.
Per aquest tractament hi ha múltiples opcions segons les diferents patents: cel·lulases alcalines, lipases, pectinases i algunes esterases. La lacasa del fongs de podridura blanca també son molt utilitzats encara que la majoria dels mètodes publicats fan servir cel·lulases i hemicel·lulases.
Les pastes de paper destintades enzimàticament tenen una qualitat superior en les propietats físiques i presenten uns nivells de tinta residual menor als tractats químicament. Variant la carga enzimàtica, temps de residència, additius i pH podem controlar el grau de destintació segons el paper que tractarem.
Patents d’enzims per “deinking”:

Eliminació del “Pitch” i de “slime”:
El “pitch” es composa bàsicament d’àcids grassos, àcids resínics, esterols, èsters de glicerol i altres grasses o ceres. És un component de la fusta soluble en metilè i suposa menys de 10% en pes de la composició però pot causar problemes en la qualitat final del paper.
Composició del pitch en diverses plantes:

L’acumulació de fangs també causa problemes a la maquinària per fabricar paper i poden ser d’origen microbià. L’acumulació de pitch es podria estalviar instaurant un medi estèril a l’interior de la fàbrica paperera però això ocasionaria grans costos i per aquest motiu no resulta rentable.
Bibliografia:
Bajpai, Pratima et al: “Enzymes improve ECF bleaching of pulp”, Thapar Centre for Indrustrial Research & Development. Patiala, India.
B. Skals, Peter (2008): “Environmental Assessment of Enzyme Assisted Processing in Pulp and Paper Industry”. Denmark. Int JLCA. 124-132.
Bajpai, Pratima (1999): “Application of Enzymes in the Pulp and Paper Industry”. Thapar Corporate Research & Development Centre. India,. Biotechnology Progress. 15(2): 147 -157.
Gutiérrez, A., J. C. del Río, et al. (2001). "The biotechnological control of pitch in paper pulp manufacturing." Trends in Biotechnology. 19(9): 340-348.
M.K. Bhat (2000): “Research review paper Cellulases and related enzymes in biotechnology”. Biotechnology Advances. 18: 355-383.
Saleem, Mahjabeen (2002): “Biobleaching of Kraft Pulp by Xylanase Produced by Bacillus
Subtilis”. International Journal of Agriculture & Biology. 4(2).
http://www.ceres.net/AboutUs/AboutUs-Biofuels-Carbo.html
http://www.thecanadianencyclopedia.com/index.cfm?PgNm=TCE&Params=A1ARTA0006564
http://aula2.elmundo.es/aula/laminas/lamina1069937674.pdf
http://www.profesorenlinea.cl/mediosocial/PapelFabricar.htm
http://www.papelnet.cl/papel/esquema_maquina.htm
http://www.lignumgroup.com/pulp
http://individual.utoronto.ca/abdel_rahman/paper/fpmp.html

El paper
El paper i els productes relacionats amb ell s’elaboren a partir de fibres de cel·lulosa presents a les plantes. Aquestes fibres poden provenir de diferents vegetals: cotó, fusta, palla de cereals, etc. Encara que la majoria de la producció de paper prové de la fusta (95%).
Existeixen diferents tipus de paper al mercat segons el tipus de fusta, acabament, ús i gramatge.
Fabricació de paper
El fonament bàsic per la fabricació de paper es trossejar el material vegetal amb aigua per formar una suspensió de fibres i després fer làmines de fibres superposades sobre una base porosa per poder filtrar l’aigua. Els papers especials com satinats i estucats es sotmeten a tractaments addicionals.
A la suspensió de fibres li diem pasta de paper o polpa que serà la matèria prima per fer els diferents tipus de paper. Existeixen dos mètodes químics d’obtenir aquesta pasta durant l’etapa de cocció: mètode alcalí o Kraft i mètode amb sulfits. Els dos treballen amb compostos del sofre però donen una pasta de paper amb propietats més adients segons el paper final que volem.
El mètode Kraft dóna una pasta marró més consistent per fer papers d’embalatge mentre que el mètode de sulfits fa una pasta de paper més dèbil i clar. Els dos processos permeten reciclatge dels productes químics però emeten gasos tòxics.
Passos en la fabricació de paper:


VÍDEOS dels pasos en la producció de paper:
La fabricación del papel
Cargado por mactiste. - Descubre más videos de ciencia y hi tech.
 Enzims implicats en la indústria del paper
Enzims implicats en la indústria del paperEs necessita una gran quantitat de processos químics per convertir els arbres en paper blanc. La indústria paperera utilitza clors per blanquejar la pasta de paper i com a resultat s’obtenen compostos orgànics clorats i subproductes amb problemes de toxicitat per l’ecosistema.
Durant el procés de cocció de la pasta de paper també s’empren sulfats que són tòxics.
Els enzims poden ajudar als fabricants de paper a reduir l’ús de productes químics perjudicials com clors i sulfats. D’aquesta manera els costos en substàncies químiques i el seu posterior tractament es redueixen de manera considerable així com l’impacte mediambiental.
Enzims com les hemicel·luloses i la xilanasa poden millorar la decoloració reduint dràsticament l’ús de clor. El tractament enzimàtic sobre la matriu de la pasta de paper permet una millor penetració dels productes químics per blanquejar i una millor extracció al rentar la lignina i compostos associats de color marró.
Composició de la fusta per fabricar paper


Millores amb enzims a la indústria paperera en cada etapa:
Blanqueig o Bleaching
L’eliminació de la lignina de la pasta química es l’etapa de “bleaching”, es necessària per raons estètiques i per la millora de les propietats del paper. Actualment per blanquejar el paper de Kraft s’utilitzen grans quantitats de clor o derivats orgànics del clor, substàncies tòxiques, mutagèniques, persistents i bio acumulatives que poden causar nombroses alteracions perjudicials als ecosistemes biològics.
Les diferents opcions de decoloració en aquest procés son: ampliar la cocció, des-lignificació per oxigen, per diòxid de clor, la utilització de peròxid de clor, clor o ozó. La majoria d’aquestes opcions, però impliquen inversions de capital i subproductes nocius. Els enzims ofereixen una reducció de l’ús del clor i altres compostos, fan que el procés sigui més senzill i s’obté un blanqueig superior en comparació amb els mètodes actuals. A més son una opció més econòmica i ecològica per la indústria del paper.

Pel bleaching enzimàtic s’han estudiat dues estratègies possibles: hemicel·luloses i enzims lignolítics.
- Hemicel·lulases: enzims per blanqueig de polpa, principalment la endo-β-xilanasa. Encara que alguns estudis demostren que enriquint les xilanases amb altres enzims hemicel·lulòsics s’incrementa l’efecte del tractament enzimàtic. Les soques per produir xilanasa a nivell comercial son: Trichoderma reesei, Thermomyces lanuginosus, Aureobasidium pullulans, y Streplividans tomyces. L’ús d’aquests enzims permet blanquejar la pasta de paper reduint un 20-25% de compostos clorats en fustes dures y un 10-15% en fustes toves. Al utilitzar xilanases la polpa s’exposa molt més al tractament químic degut a la degradació de la hemicel·lulosa.
- Enzims lignolítics: ataquen directament la lignina i és més eficaç. Els fongs de la pudrició blanca (Pleurotus, Coriolus, Phlebia, Poria i Ceriporiopsis) són els que produeixen els enzims lignin-peroxidasa, i lacases.

Modificació de fibres o fibrilació:
Modificant fibres s’aconsegueix disminuir el consum d’energia a la fase tèrmica i mecànica de la producció de polpa. El tractament enzimàtic es produeix a la superfície externa de la fibra, dissolent una petita quantitat de fibra i augmentant la hidratació.
Si es combina amb un pretractament amb agents alcalins, l’acció enzimàtica redueix de manera important l’energia al procés mecànic per refinar la polpa de paper.
Els microorganismes per obtenir els enzims necessaris son Bacillus i algunes espècies d’Aspergillus.
Solubilització de la pasta:
Pastes que es dissolen per produir materials cel·lulòsics com acetats, cel·lofanes i raions. Es fa derivatitzant i solubilitzant amb alta puresa, encara no hi ha enzims específics.
Destintació o “Deinking”:
La destintació enzimàtica converteix les fibres secundàries (fibres disgregades de la pasta de paper) en productes de qualitat. Resulta un mètode més econòmic i eficaç per tractar residus de paper de reciclatge.
Per aquest tractament hi ha múltiples opcions segons les diferents patents: cel·lulases alcalines, lipases, pectinases i algunes esterases. La lacasa del fongs de podridura blanca també son molt utilitzats encara que la majoria dels mètodes publicats fan servir cel·lulases i hemicel·lulases.
Les pastes de paper destintades enzimàticament tenen una qualitat superior en les propietats físiques i presenten uns nivells de tinta residual menor als tractats químicament. Variant la carga enzimàtica, temps de residència, additius i pH podem controlar el grau de destintació segons el paper que tractarem.
Patents d’enzims per “deinking”:

Eliminació del “Pitch” i de “slime”:
El “pitch” es composa bàsicament d’àcids grassos, àcids resínics, esterols, èsters de glicerol i altres grasses o ceres. És un component de la fusta soluble en metilè i suposa menys de 10% en pes de la composició però pot causar problemes en la qualitat final del paper.
Composició del pitch en diverses plantes:

L’acumulació de fangs també causa problemes a la maquinària per fabricar paper i poden ser d’origen microbià. L’acumulació de pitch es podria estalviar instaurant un medi estèril a l’interior de la fàbrica paperera però això ocasionaria grans costos i per aquest motiu no resulta rentable.
Bibliografia:
Bajpai, Pratima et al: “Enzymes improve ECF bleaching of pulp”, Thapar Centre for Indrustrial Research & Development. Patiala, India.
B. Skals, Peter (2008): “Environmental Assessment of Enzyme Assisted Processing in Pulp and Paper Industry”. Denmark. Int JLCA. 124-132.
Bajpai, Pratima (1999): “Application of Enzymes in the Pulp and Paper Industry”. Thapar Corporate Research & Development Centre. India,. Biotechnology Progress. 15(2): 147 -157.
Gutiérrez, A., J. C. del Río, et al. (2001). "The biotechnological control of pitch in paper pulp manufacturing." Trends in Biotechnology. 19(9): 340-348.
M.K. Bhat (2000): “Research review paper Cellulases and related enzymes in biotechnology”. Biotechnology Advances. 18: 355-383.
Saleem, Mahjabeen (2002): “Biobleaching of Kraft Pulp by Xylanase Produced by Bacillus
Subtilis”. International Journal of Agriculture & Biology. 4(2).
http://www.ceres.net/AboutUs/AboutUs-Biofuels-Carbo.html
http://www.thecanadianencyclopedia.com/index.cfm?PgNm=TCE&Params=A1ARTA0006564
http://aula2.elmundo.es/aula/laminas/lamina1069937674.pdf
http://www.profesorenlinea.cl/mediosocial/PapelFabricar.htm
http://www.papelnet.cl/papel/esquema_maquina.htm
http://www.lignumgroup.com/pulp
http://individual.utoronto.ca/abdel_rahman/paper/fpmp.html