Diana Navarro i Ana Rodriguez. Curs 2009-10
Objetivo
Mejorar la actividad hidrolítica de la glucoamilasa y su capacidad desramificante sobre los enlaces α-1,6 de la molécula del almidón por técnicas de mutagénesis dirigida y aleatoria. Para ello se construyen diferentes plásmidos.
Introducción
La glucoamilasa es una enzima hidrolítica del grupo de las amilasas, también conocida como amiloglucosidasa, su nombre sistemático es 1,4-alfa-D-glucano glucohidrolasa (EC 3.2.1.3). Es una de las enzimas más estudiadas debido a su influencia directa en la degradación del almidón, uno de los productos alimentarios más explotados a nivel mundial.
Las cadenas de almidón están compuestas por dos grandes sub-cadenas, amilosa y amilopectina. La función de la glucoamilasa es actuar en la reacción de hidrólisis en cadenas de polisacáridos rompiendo los enlaces 1,4-alfa-D-glucosa que se encuentran de manera residual en las cadenas después de haber sido digeridas por alfa y beta amilasas. El principal producto final de la acción de la glucoamilasa sobre el almidón es glucosa, lo que la diferencia de la alfa y beta amilasas.
Su actividad es máxima entre pH 4 - 5,5 y temperatura alrededor de 55ºC - 65ºC, es producida extracelularmente por numerosos tipos de hongos y algunas bacterias, aunque la principal fuente de esta enzima son los hongos filamentosos donde destaca el género Aspergillus. También se han utilizado algunos cultivos de bacterias (Bacillus coagulans y Lactobacillus brevis) y levaduras (Saccharomyces cerevisiae y Sacharomycopsis figuligera) aunque la baja producción dificulta su comercialización.
Aplicaciones de la glucoamilasa
Interviene en los pasos de sacarificación de:
Las amilasas han sido de gran importancia en diversas industrias durante las últimas décadas debido a la utilización de las técnicas biotecnológicas basadas en enzimas, optimizando así el proceso de degradación del almidón. Las amilasas ocupan cerca de un 25% en el mercado enzimático llegando a sustituir completamente procesos de hidrólisis química en la industria del almidón debido fundamentalmente a su termoestabilidad y a que la hidrólisis se lleva a cabo en un único paso.
Proceso de hidrólisis
La hidrólisis del enlace glicosídico tiene lugar mediante una catálisis ácida que requiere la actuación de dos residuos localizados en el centro activo del enzima, en la mayor parte de los casos aspártico o glutámico. Uno de ellos actúa como donador de un protón y el otro como nucleófilo/base. Esta hidrólisis puede ocurrir a través de dos mecanismos diferentes que dan como resultado la retención o la inversión de la configuración del carbono anomérico. La disposición espacial y distancia entre los residuos catalíticos es diferente según sea el mecanismo de hidrólisis del enlace.
Genes de la glucoamilasa
Saccharomyces cerevisiae presenta tres genes distintos, no ligados, STA1, STA2 y STA3 que codifican formas idénticas de glucoamilasa denominadas GAI, GAII y GAIII. Además de la glucoamilasa codificada por estos genes STA, existe otra forma de la enzima codificada por el gen SGA1 (Sporulationspecific glucoamylase), producida durante los procesos de meiosis y esporulación, la cual es responsable de la degradación del glucógeno intracelular. El péptido señal necesario para la secreción de la enzima se encuentra en la zona 5’ de los genes STA la cual es una zona rica en serina y treonina.
Estructura tridimensional de la glucoamilasa
La mayoría de las glucoamilasas son enzimas multidominio, es decir, están constituidas por un dominio catalítico (CD, catalytic domain) unido, mediante una región glicosilada de longitud variable, a un dominio de unión al almidón (SBD, starch binding domain). La glucoamilasa de S.cerevisiae se caracteriza por poseer un dominio rico en serina y treonina en lugar de un dominio de unión al almidón en su región amino terminal.
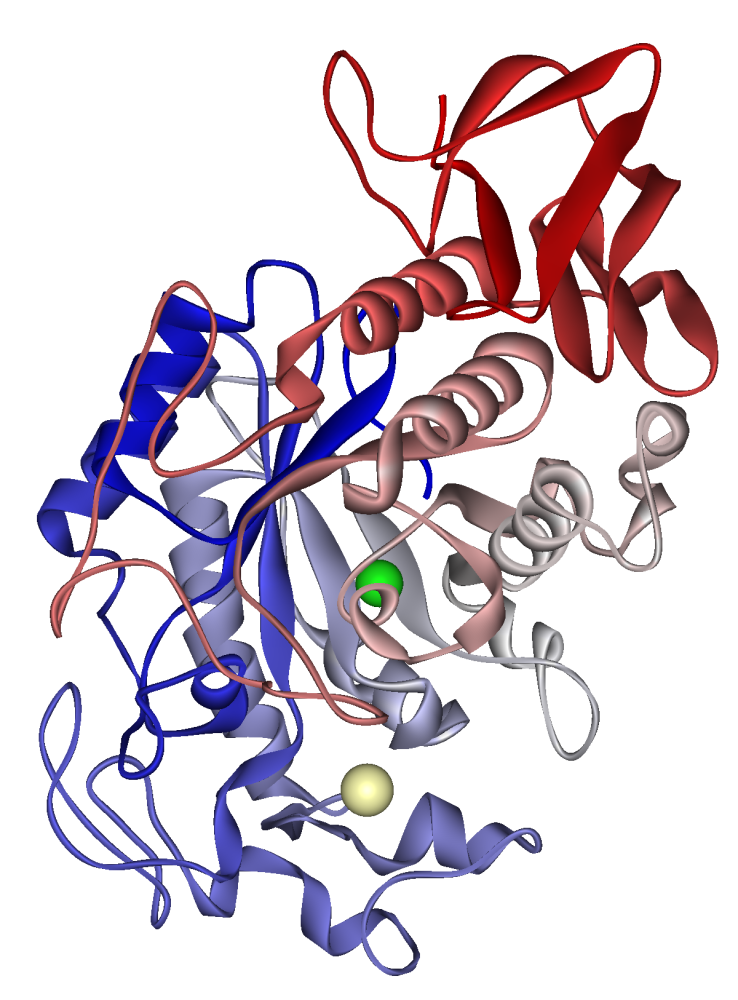
La mayoría de los dominios catalíticos de las glucoamilasas presentan una arquitectura similar que comprende como mínimo doce hélices alfa, apareadas dos a dos formando un barril (α/α)6. En el CD existen regiones muy conservadas de la secuencia que comprenden bucles o loops que rodean al centro activo. Estos bucles, que carecen de estructura secundaria definida, se han relacionado con la estabilidad y especificidad de substrato.
Una característica muy importante, desde el punto de vista funcional de las glucoamilasas de levadura es la ausencia del SBD. La falta de este dominio no reduce la eficacia de los enzimas cuando actúan sobre el almidón soluble y varios estudios han confirmado la importancia de este elemento en la degradación de moléculas grandes de almidón insoluble.
Las glucoamilasas presentan múltiples sitios de N-glicosilación pero sólo tres aparecen conservados en distintas subfamilias. Estos están localizados en los loops del CD de la cadena polipeptídica desprovistos de estructura secundaria. Los carbohidratos asociados a glucoamilasas parecen tener un efecto estabilizador que además dirigen el correcto plegamiento de la proteína.
Determinación de la actividad de la glucoamilasa.
Para saber si realmente hay cambios significativos respecto a la glucoamilasa nativa y las diferentes variedades mutantes se ha de determinar su actividad. El método semicuantitativo consiste en observar halos de hidrólisis alrededor de las colonias transformadas sembradas en placas con 1% de almidón insoluble en el medio. El método cuantitativo consiste en medir la cantidad de glucosa liberada a partir de alimón soluble al 0,5%, almidón insoluble o pululano al 1%.
Al final, todas las estirpes transformadas con el plásmido pS2 presentan actividad glucoamilasa.
Esta actividad glucoamilasa detectada en las estirpes estudiadas es un reflejo de los niveles de expresión del gen STA1, de forma que a mayor expresión del gen, mayor actividad glucoamilasa.
Una vez determinada la actividad del enzima se procede a generar diversos mutantes.
Modificación estructural de la glucoamilasa de S.cerevisiae para mejorar su actividad catalítica
La ingeniería de proteínas y más en concreto la evolución dirigida permite la mejora de enzimas con aplicación industrial. Este proceso, que mimetiza la evolución natural, se lleva a cabo mediante etapas alternas de generación de proteínas mutantes, con otras de selección de clones con propiedades mejoradas. Las mutaciones se introducen en la secuencia de nucleótidos tanto en lugares específicos mediante mutagénesis dirigida (denominando el proceso de "diseño racional") como al azar por mutagénesis aleatoria.
La mejora de las propiedades, que en nuestro caso es la actividad catalítica de la glucoamilasa de S. cerevisiae, tiene interés biotecnológico debido a su posible aplicación en la bioconversión de almidón en etanol y en otros productos de fermentación. Esto se realiza mediante la aplicación de técnicas de mutagénesis dirigida y aleatoria del gen STA1.
Mutagénesis dirigida
La mutagénesis dirigida consiste en amplificar la secuencia completa de un plásmido portador del gen o fragmento que se pretende mutagenizar con dos oligonucleótidos divergentes, cuyos puntos de inicio están situados en posiciones nucleotídicas adyacentes, uno de los cuales contiene la mutación a introducir.
Como ya hemos dicho antes, el dominio catalítico de las glucoamilasas contiene regiones denominadas bucles o loops. Mediante esta técnica se modifican los residuos de las regiones de los loops 3 y 5 de la glucoamilasa (GA) de S. cerevisiae, con el objetivo de mimetizar la estructura de la GA de H. resinae con capacidad desramificante. La información relativa a otras glucoamilasas de la misma familia, indica que estas regiones tienen influencia en la especificidad sobre los enlaces α-1,6 de la molécula de almidón.
En los mutante L3 y L5 se modificaron los loops 3 o 5 por separado, realizando los cambios en base a la secuencia de la GA de H. resinae, el mutante L3L5 combina los dos grupos de cambios y en el mutante LD5 se modificó el loop 5 eliminando una región no conservada de cinco aminoácidos que está presente en S. cerevisiae.
Esta aproximación, por “diseño racional”, no proporcionó resultados positivos, ya que ninguna de las variantes obtenidas posee actividad desramificante y en la mayoría de los casos se perdió la capacidad de hidrolizar enlaces α-1,4.
Probablemente, las modificaciones en la secuencia primaria causan distorsiones estructurales que impiden la producción de un enzima funcional. Por otra parte, los conocimientos actuales sobre las relaciones estructura-función en proteínas suelen ser insuficientes para hacer del diseño racional una aproximación viable en los procesos de evolución dirigida.
La dificultad es el introducir cambios en residuos próximos al centro catalítico sin causar modificaciones no deseadas que afecten a la actividad, modificaciones difícilmente predecibles en base al modelo estructural. En concreto, el loop3 se encuentra muy próximo al residuo catalítico E521 y cualquier modificación en este entorno es previsible que afecte la configuración del sitio catalítico. En el loop 5 de la región catalítica de S. cerevisiae se encuentra presente un segmento de 5 residuos, no presente en otras glucoamilasas de la familia, que puede estorbar el acceso del substrato al centro catalítico y dificultar, por tanto, la hidrólisis de los enlaces α-1,6.
Mutagénesis aleatoria.
Los experimentos de mutagénesis aleatoria permiten encontrar caminos para la mejora enzimática que no pueden ser anticipados por un diseño racional. En este caso la mutagénesis aleatoria del gen STA1 se realizó por error prone-PCR y recombinación de DNA in vitro (también denominado DNAshuffling). Se ensayaron aproximadamente unos 10.000 transformantes, obtenidos en 3 experimentos independientes de mutagénesis aleatoria.
La mutagénesis por error prone-PCR se inició utilizando como DNA molde el plásmido pYS2 (plásmido pYES2 con un inserto SacI/XbaI de 2.5 kb) que contiene STA1. El gen se amplificó utilizando como cebadores los oligonucleótidos LL481 y LL482. Se llevaron a cabo cuatro reacciones paralelas de PCR, cada una de las cuales contenía un dNTP en exceso frente a los demás dNTPs que fueron utilizados a la concentración estándar.
Aplicando la técnica de DNA–shuffling al gen STA1 de S.cerevisiae y a versiones del mismo (L3, L5, L35, L35D) obtenidas por mutagénesis dirigida se obtienen diferentes versiones mutantes con las que se construye una biblioteca de mutantes.
Tras un proceso de screening o búsqueda de mutantes con mayor actividad glucoamilasa se selecciona la versión con mayor actividad, designada Cl24p (GAproducida por la estirpe Cl24 que presenta elevada actividad). Su secuenciación muestra que la proteína mutante tiene tres mutaciones puntuales y una deleción parcial. Esta secuenciación se lleva a cabo para saber qué cambios son los que han incrementado su actividad.
Los cambios en la secuencia nucleotídica de Cl24p se muestran en la Figura 6 donde aparecen listados los nucleótidos que difieren, las alteraciones que se producen tanto en el gen como en la proteína y la región donde se encuentra el cambio dentro de los dominios de la glucoamilasa de S. cerevisiae.
La secuenciación muestra nueve cambios en la secuencia nucleotídica de la versión mutante del gen STA1 aunque únicamente cuatro de ellos producen alteraciones en la proteína Cl24p con respecto a la glucoamilasa nativa. En el resto de los cambios, las alteraciones de la secuencia de nucleótidos no se traducen en la secuencia proteica. La Figura 7 muestra los residuos reemplazados en la secuencia primaria de la glucoamilasa. Como puede observarse, los cambios se han producido en la región rica en serina y treonina sin afectar en ningún caso al dominio catalítico.
MVGLKNPYTHTMQRPFLLAYLVLSLLFNSALGFPTALVPRGSSSSNITSSGPSSTPFSSATESFSTGTTVTPSSSKYPGSKTETSVSSTTETTIVPTTTTTSVITPST[TTITTTVCSTGTNSAGETTSGCSPKTITTTVPCSTSPSETASESTTTSPTTPVTTVVSTTVVTTEYSTSTKQGGEITTTFVTKNIPTTYLTTIAPTSSVTTVTNFTP]TTITTTVCSTGTNSAGETTSGCSPKTVTTTVPCSTGTGEYTTEATAPVTTAVTTTVVTTESSTGTNSAGKTTTSYTTKSVPTTYVFDFGKGILDQSCGGFSNNGSSQVQLRDVVLMNGTVVYDSNGAWDSSALEEWLQRQKKVSIERIFENIGPSAVYPSILPGVVIASPSQTHPDYFYQWIRDSALTINSIVSHSADPAIETLLQYLNVSFHLQRTNNTLGAGIGYTNDTVALGDPKWNVDNTAFTEPWGRPQNDGPALRSIAILKIIDYIKQSGTDLGAKYPFQSTADIFDDIVRWDLRFIIDHWNSSGFDLWEEVNGMHFFTLLVQLSAVDRSLSYFNASERSSPFVEELRQTRRDISKFLVDPANGFINGKYNYIVETPMIADTLRSGLDISTLLAANTVHDAPSASHLPFDINDPAVLNTLHHLMLHMRSIYPINDSSKNATGIALGRYPEDVYDGYGVGEGNPWVLATCAASTTLYQLIYRHISEQHDLVVPMNNDCSNAFWSELVFSNLTTLGNDEGYLILEFNTPAFNQTIQKIFQLADSFLGQAESHVGTDGELSEQFNKYTGFMQGAQHLTWSYTSFWDAYQIRQEVLQSL
Estructura primaria de la glucoamilasa de S. cerevisiae. En negrita se señala el péptido señal; en rosa el dominio rico en serina y treonina; y en azul el dominio catalítico. Los cambios existentes en la forma mutante del enzima se indican en sombreado negro y son S74T, T286M y Q354L. La secuencia de 107 aminoácidos pérdida, desde T109 hasta P215 ambos inclusive, se indica entre corchetes en negrita.
Los cambios Ser74Thr, Thr286Met y Gln354Leu parecen poco significativos porque se localizan en la superficie de la proteína, zona que no es previsible que afecte de forma significativa a su estructura tridimensional.
El cambio más notable es la deleción de 107 aminoácidos producida al principio de la región rica en serina y treonina, en concreto, la presencia de una secuencia palindrómica, TTITT que aparece en la zona adyacente a la deleción. Este cambio, como se observa en el modelo, elimina uno de los tres barriles beta que forman la región rica en serina y treonina, en concreto el primero, y altera la estructura secundaria del segundo.
Conclusiones
En ninguna de las variantes de la glucoamilasa de S. cerevisiae se obtuvo una mejora de la actividad frente a enlaces α-1,6. Sin embargo, el proceso de mutagénesis aleatoria permitió obtener una versión más activa que el enzima nativo.
Los modelos estructurales del dominio catalítico y de la región rica en Ser/Thr de la glucoamilasa de S. cerevisiae explican muchas propiedades del enzima y nos han permitido diseñar nuevas versiones con el objeto de mejorar su actividad.
La mutagénesis aleatoria por el método de DNA-Shuffling ha permitido obtener una variante de glucoamilasa de S. cerevisiae con actividad mejorada respecto al enzima nativo. El estudio de la secuencia del mutante indica que la mejora podría deberse a una deleción parcial de la región amino terminal de STA1. Este estudio se ha realizado mediante la construcción de una versión modificada de Sta1p (denominada Sta1dp), en la que se ha delecionado la mayor parte de la región rica en Ser/Thr de la proteína, presenta actividad glucoamilasa. Por lo que se puede concluir que la región rica en Ser/Thrno es crítica para la actividad. El éxito de esta aproximación sugiere la posibilidad de nuevas mejoras por evolución dirigida de las versiones más activas del enzima.
BIBLIOGRAFÍA
Objetivo
Mejorar la actividad hidrolítica de la glucoamilasa y su capacidad desramificante sobre los enlaces α-1,6 de la molécula del almidón por técnicas de mutagénesis dirigida y aleatoria. Para ello se construyen diferentes plásmidos.
Introducción
La glucoamilasa es una enzima hidrolítica del grupo de las amilasas, también conocida como amiloglucosidasa, su nombre sistemático es 1,4-alfa-D-glucano glucohidrolasa (EC 3.2.1.3). Es una de las enzimas más estudiadas debido a su influencia directa en la degradación del almidón, uno de los productos alimentarios más explotados a nivel mundial.
Las cadenas de almidón están compuestas por dos grandes sub-cadenas, amilosa y amilopectina. La función de la glucoamilasa es actuar en la reacción de hidrólisis en cadenas de polisacáridos rompiendo los enlaces 1,4-alfa-D-glucosa que se encuentran de manera residual en las cadenas después de haber sido digeridas por alfa y beta amilasas. El principal producto final de la acción de la glucoamilasa sobre el almidón es glucosa, lo que la diferencia de la alfa y beta amilasas.
Su actividad es máxima entre pH 4 - 5,5 y temperatura alrededor de 55ºC - 65ºC, es producida extracelularmente por numerosos tipos de hongos y algunas bacterias, aunque la principal fuente de esta enzima son los hongos filamentosos donde destaca el género Aspergillus. También se han utilizado algunos cultivos de bacterias (Bacillus coagulans y Lactobacillus brevis) y levaduras (Saccharomyces cerevisiae y Sacharomycopsis figuligera) aunque la baja producción dificulta su comercialización.
Aplicaciones de la glucoamilasa
Interviene en los pasos de sacarificación de:
- Producción de energía: se utiliza para la producción de etanol.
- Conversión enzimática del almidón: se utiliza junto con la alfa amilasa para la conversión industrial del almidón a glucosa.
Las amilasas han sido de gran importancia en diversas industrias durante las últimas décadas debido a la utilización de las técnicas biotecnológicas basadas en enzimas, optimizando así el proceso de degradación del almidón. Las amilasas ocupan cerca de un 25% en el mercado enzimático llegando a sustituir completamente procesos de hidrólisis química en la industria del almidón debido fundamentalmente a su termoestabilidad y a que la hidrólisis se lleva a cabo en un único paso.
Proceso de hidrólisis
La hidrólisis del enlace glicosídico tiene lugar mediante una catálisis ácida que requiere la actuación de dos residuos localizados en el centro activo del enzima, en la mayor parte de los casos aspártico o glutámico. Uno de ellos actúa como donador de un protón y el otro como nucleófilo/base. Esta hidrólisis puede ocurrir a través de dos mecanismos diferentes que dan como resultado la retención o la inversión de la configuración del carbono anomérico. La disposición espacial y distancia entre los residuos catalíticos es diferente según sea el mecanismo de hidrólisis del enlace.
Genes de la glucoamilasa
Saccharomyces cerevisiae presenta tres genes distintos, no ligados, STA1, STA2 y STA3 que codifican formas idénticas de glucoamilasa denominadas GAI, GAII y GAIII. Además de la glucoamilasa codificada por estos genes STA, existe otra forma de la enzima codificada por el gen SGA1 (Sporulationspecific glucoamylase), producida durante los procesos de meiosis y esporulación, la cual es responsable de la degradación del glucógeno intracelular. El péptido señal necesario para la secreción de la enzima se encuentra en la zona 5’ de los genes STA la cual es una zona rica en serina y treonina.
Estructura tridimensional de la glucoamilasa
La mayoría de las glucoamilasas son enzimas multidominio, es decir, están constituidas por un dominio catalítico (CD, catalytic domain) unido, mediante una región glicosilada de longitud variable, a un dominio de unión al almidón (SBD, starch binding domain). La glucoamilasa de S.cerevisiae se caracteriza por poseer un dominio rico en serina y treonina en lugar de un dominio de unión al almidón en su región amino terminal.
La mayoría de los dominios catalíticos de las glucoamilasas presentan una arquitectura similar que comprende como mínimo doce hélices alfa, apareadas dos a dos formando un barril (α/α)6. En el CD existen regiones muy conservadas de la secuencia que comprenden bucles o loops que rodean al centro activo. Estos bucles, que carecen de estructura secundaria definida, se han relacionado con la estabilidad y especificidad de substrato.
Una característica muy importante, desde el punto de vista funcional de las glucoamilasas de levadura es la ausencia del SBD. La falta de este dominio no reduce la eficacia de los enzimas cuando actúan sobre el almidón soluble y varios estudios han confirmado la importancia de este elemento en la degradación de moléculas grandes de almidón insoluble.
Las glucoamilasas presentan múltiples sitios de N-glicosilación pero sólo tres aparecen conservados en distintas subfamilias. Estos están localizados en los loops del CD de la cadena polipeptídica desprovistos de estructura secundaria. Los carbohidratos asociados a glucoamilasas parecen tener un efecto estabilizador que además dirigen el correcto plegamiento de la proteína.
Determinación de la actividad de la glucoamilasa.
Para saber si realmente hay cambios significativos respecto a la glucoamilasa nativa y las diferentes variedades mutantes se ha de determinar su actividad. El método semicuantitativo consiste en observar halos de hidrólisis alrededor de las colonias transformadas sembradas en placas con 1% de almidón insoluble en el medio. El método cuantitativo consiste en medir la cantidad de glucosa liberada a partir de alimón soluble al 0,5%, almidón insoluble o pululano al 1%.
Al final, todas las estirpes transformadas con el plásmido pS2 presentan actividad glucoamilasa.
Esta actividad glucoamilasa detectada en las estirpes estudiadas es un reflejo de los niveles de expresión del gen STA1, de forma que a mayor expresión del gen, mayor actividad glucoamilasa.
Una vez determinada la actividad del enzima se procede a generar diversos mutantes.
Modificación estructural de la glucoamilasa de S.cerevisiae para mejorar su actividad catalítica
La ingeniería de proteínas y más en concreto la evolución dirigida permite la mejora de enzimas con aplicación industrial. Este proceso, que mimetiza la evolución natural, se lleva a cabo mediante etapas alternas de generación de proteínas mutantes, con otras de selección de clones con propiedades mejoradas. Las mutaciones se introducen en la secuencia de nucleótidos tanto en lugares específicos mediante mutagénesis dirigida (denominando el proceso de "diseño racional") como al azar por mutagénesis aleatoria.
La mejora de las propiedades, que en nuestro caso es la actividad catalítica de la glucoamilasa de S. cerevisiae, tiene interés biotecnológico debido a su posible aplicación en la bioconversión de almidón en etanol y en otros productos de fermentación. Esto se realiza mediante la aplicación de técnicas de mutagénesis dirigida y aleatoria del gen STA1.
Mutagénesis dirigida
La mutagénesis dirigida consiste en amplificar la secuencia completa de un plásmido portador del gen o fragmento que se pretende mutagenizar con dos oligonucleótidos divergentes, cuyos puntos de inicio están situados en posiciones nucleotídicas adyacentes, uno de los cuales contiene la mutación a introducir.
Como ya hemos dicho antes, el dominio catalítico de las glucoamilasas contiene regiones denominadas bucles o loops. Mediante esta técnica se modifican los residuos de las regiones de los loops 3 y 5 de la glucoamilasa (GA) de S. cerevisiae, con el objetivo de mimetizar la estructura de la GA de H. resinae con capacidad desramificante. La información relativa a otras glucoamilasas de la misma familia, indica que estas regiones tienen influencia en la especificidad sobre los enlaces α-1,6 de la molécula de almidón.
En los mutante L3 y L5 se modificaron los loops 3 o 5 por separado, realizando los cambios en base a la secuencia de la GA de H. resinae, el mutante L3L5 combina los dos grupos de cambios y en el mutante LD5 se modificó el loop 5 eliminando una región no conservada de cinco aminoácidos que está presente en S. cerevisiae.
Esta aproximación, por “diseño racional”, no proporcionó resultados positivos, ya que ninguna de las variantes obtenidas posee actividad desramificante y en la mayoría de los casos se perdió la capacidad de hidrolizar enlaces α-1,4.
Probablemente, las modificaciones en la secuencia primaria causan distorsiones estructurales que impiden la producción de un enzima funcional. Por otra parte, los conocimientos actuales sobre las relaciones estructura-función en proteínas suelen ser insuficientes para hacer del diseño racional una aproximación viable en los procesos de evolución dirigida.
La dificultad es el introducir cambios en residuos próximos al centro catalítico sin causar modificaciones no deseadas que afecten a la actividad, modificaciones difícilmente predecibles en base al modelo estructural. En concreto, el loop3 se encuentra muy próximo al residuo catalítico E521 y cualquier modificación en este entorno es previsible que afecte la configuración del sitio catalítico. En el loop 5 de la región catalítica de S. cerevisiae se encuentra presente un segmento de 5 residuos, no presente en otras glucoamilasas de la familia, que puede estorbar el acceso del substrato al centro catalítico y dificultar, por tanto, la hidrólisis de los enlaces α-1,6.
Mutagénesis aleatoria.
Los experimentos de mutagénesis aleatoria permiten encontrar caminos para la mejora enzimática que no pueden ser anticipados por un diseño racional. En este caso la mutagénesis aleatoria del gen STA1 se realizó por error prone-PCR y recombinación de DNA in vitro (también denominado DNAshuffling). Se ensayaron aproximadamente unos 10.000 transformantes, obtenidos en 3 experimentos independientes de mutagénesis aleatoria.
La mutagénesis por error prone-PCR se inició utilizando como DNA molde el plásmido pYS2 (plásmido pYES2 con un inserto SacI/XbaI de 2.5 kb) que contiene STA1. El gen se amplificó utilizando como cebadores los oligonucleótidos LL481 y LL482. Se llevaron a cabo cuatro reacciones paralelas de PCR, cada una de las cuales contenía un dNTP en exceso frente a los demás dNTPs que fueron utilizados a la concentración estándar.
Aplicando la técnica de DNA–shuffling al gen STA1 de S.cerevisiae y a versiones del mismo (L3, L5, L35, L35D) obtenidas por mutagénesis dirigida se obtienen diferentes versiones mutantes con las que se construye una biblioteca de mutantes.
Tras un proceso de screening o búsqueda de mutantes con mayor actividad glucoamilasa se selecciona la versión con mayor actividad, designada Cl24p (GAproducida por la estirpe Cl24 que presenta elevada actividad). Su secuenciación muestra que la proteína mutante tiene tres mutaciones puntuales y una deleción parcial. Esta secuenciación se lleva a cabo para saber qué cambios son los que han incrementado su actividad.
Los cambios en la secuencia nucleotídica de Cl24p se muestran en la Figura 6 donde aparecen listados los nucleótidos que difieren, las alteraciones que se producen tanto en el gen como en la proteína y la región donde se encuentra el cambio dentro de los dominios de la glucoamilasa de S. cerevisiae.
La secuenciación muestra nueve cambios en la secuencia nucleotídica de la versión mutante del gen STA1 aunque únicamente cuatro de ellos producen alteraciones en la proteína Cl24p con respecto a la glucoamilasa nativa. En el resto de los cambios, las alteraciones de la secuencia de nucleótidos no se traducen en la secuencia proteica. La Figura 7 muestra los residuos reemplazados en la secuencia primaria de la glucoamilasa. Como puede observarse, los cambios se han producido en la región rica en serina y treonina sin afectar en ningún caso al dominio catalítico.
Estructura primaria de la glucoamilasa de S. cerevisiae. En negrita se señala el péptido señal; en rosa el dominio rico en serina y treonina; y en azul el dominio catalítico. Los cambios existentes en la forma mutante del enzima se indican en sombreado negro y son S74T, T286M y Q354L. La secuencia de 107 aminoácidos pérdida, desde T109 hasta P215 ambos inclusive, se indica entre corchetes en negrita.
Los cambios Ser74Thr, Thr286Met y Gln354Leu parecen poco significativos porque se localizan en la superficie de la proteína, zona que no es previsible que afecte de forma significativa a su estructura tridimensional.
El cambio más notable es la deleción de 107 aminoácidos producida al principio de la región rica en serina y treonina, en concreto, la presencia de una secuencia palindrómica, TTITT que aparece en la zona adyacente a la deleción. Este cambio, como se observa en el modelo, elimina uno de los tres barriles beta que forman la región rica en serina y treonina, en concreto el primero, y altera la estructura secundaria del segundo.
Conclusiones
En ninguna de las variantes de la glucoamilasa de S. cerevisiae se obtuvo una mejora de la actividad frente a enlaces α-1,6. Sin embargo, el proceso de mutagénesis aleatoria permitió obtener una versión más activa que el enzima nativo.
Los modelos estructurales del dominio catalítico y de la región rica en Ser/Thr de la glucoamilasa de S. cerevisiae explican muchas propiedades del enzima y nos han permitido diseñar nuevas versiones con el objeto de mejorar su actividad.
La mutagénesis aleatoria por el método de DNA-Shuffling ha permitido obtener una variante de glucoamilasa de S. cerevisiae con actividad mejorada respecto al enzima nativo. El estudio de la secuencia del mutante indica que la mejora podría deberse a una deleción parcial de la región amino terminal de STA1. Este estudio se ha realizado mediante la construcción de una versión modificada de Sta1p (denominada Sta1dp), en la que se ha delecionado la mayor parte de la región rica en Ser/Thr de la proteína, presenta actividad glucoamilasa. Por lo que se puede concluir que la región rica en Ser/Thrno es crítica para la actividad. El éxito de esta aproximación sugiere la posibilidad de nuevas mejoras por evolución dirigida de las versiones más activas del enzima.
BIBLIOGRAFÍA
- CALVO, Miguel. “Estructura del almidón” http://milksci.unizar.es/bioquimica/temas/azucares/almidon.html [Consulta 15 Ocutbre 2009]
- CEDANO, Juan Antonio. “Rediseño de la eficiencia catalítica y de la termoresistencia de la beta(1-3)(1-4)glucanasa de Bacillus licheniformis. Institut de Biotecnologia i Biomedicina, Universitat Autònoma de Barcelona.
- CHAPLIN, Martin. “The use of enzymes in starch hydrolysis” http://www1.lsbu.ac.uk/biology/enztech/starch.html [Consulta 20 Octubre 2009]
- E. OPHARDT, Charles. “Starch”, http://www.elmhurst.edu/~chm/vchembook/547starch.html [Consulta 23 Octubre 2009]
- ELIÉCER CARRERA, JORGE. Producción y Aplicación de enzimas industriales. Facultad de ciencias agropecuarias, Vol 1, No1, Marzo 2003.
- LATORRE GARCÍA, LORENA. Análisis Estructural y Modificación Funcional de la Glucoamilasa de Saccharomyces cerevisiae. Departamento de Bioquímica y Biología molecular. Universidad de Valéncia, 2008.
- Practical Biotechnology. “Glucoamylase production by yeast”. http://www.ncbe.reading.ac.uk/ncbe/protocols/PRACBIOTECH/PDF/amgprod.pdf [Consulta 20 Octubre 2009]
- http://www.chem.qmul.ac.uk/iubmb/enzyme/EC3/2/1/3.html [Consulta 15 Octubre 2009]
- http://www.pdb.org/pdb/results/results.do?gotopage=1 [Consulta 15 Ocutbre 2009]